How Does the Brain Create Social Network Maps?
Post by Amanda Engstrom
The takeaway
The brain constructs an internal map of social networks, even for relationships never directly observed. New research shows that the hippocampus and entorhinal cortex encode these maps, enabling us to infer and navigate complex real-world social connections.
What's the science?
The brain's ability to navigate complex social networks, like tracking how individuals are connected within a broader group, is fundamental to human social life. Neurons in the medial temporal lobe (MTL) are known to encode cognitive maps of physical space, and converging evidence suggests the MTL may similarly represent abstract relational structures; however, how the brain encodes large-scale, real-world social networks remains unknown. This week in PNAS, Teoh and colleagues combined computational modeling with fMRI and a longitudinal study of a real-world social network to investigate how activity in the hippocampus and entorhinal cortex scales to represent naturalistic social structures.
How did they do it?
To uncover how social network structure is encoded across individuals, the authors recruited undergraduates within a real-world social network and asked them to identify their friends within that network at multiple timepoints, generating a ground-truth friendship map (defined as mutual recognition between two individuals). Participants were asked to judge the relationships between other network members in a pairwise manner, including pairs they had never directly interacted with, to assess participants' ability to infer unseen social connections. The authors then tested four computational models of network representation to determine which best explained participants' behavioral patterns. These models varied in their relational complexity, the weight they gave to connections between individuals, the shortest path distance between two people, and the structural complexity of those paths.
To identify the brain regions supporting these representations, the authors used functional MRI (fMRI) to measure MTL activity while participants viewed photographs of community members and judged whether each person belonged to their network. Regions of interest included both the left and right hemispheres of the anterior hippocampus (aHC), posterior hippocampus (pHC), and the entorhinal cortex (EC). To link brain activity patterns to the computational models, they applied representational similarity analysis (RSA), a method that asks whether the pattern of similarity across brain responses mirrors the structure predicted by a given model of representation.
Finally, to assess whether these neural maps translate into functional social reasoning, the authors administered an Information Flow task where participants were asked to determine how information would travel from one participant across the network; a measure of how well participants could navigate their social network to trace indirect connections. Responses were linked to individual neural representations to determine how MTL activity patterns relate to real-world social inference.
What did they find?
Participants demonstrated structured, non-random judgments about unseen relationships, and their accuracy decreased with social distance: participants were most accurate when judging pairs close to their own friendships, with performance declining as the path distance between pairs increased. This suggests that participants do not simply recall direct ties but actively infer indirect connections using a structured internal representation of the network. Of the computational models tested, the Katz communicability model, which captures integration across multiple indirect paths between individuals, rather than just the shortest or most direct route, provided the best fit to participants' behavioral data. This indicates that people represent their social networks as distributed, multi-path structures rather than simple maps of direct connections.
At the neural level, the RSA revealed that the right EC encoded an abstract map of multistep network connections, while the right aHC may encode a more veridical representation of directly observed social ties. This regional dissociation suggests the MTL supports social network representation through at least two complementary encoding mechanisms, paralleling its known role in spatial navigation.
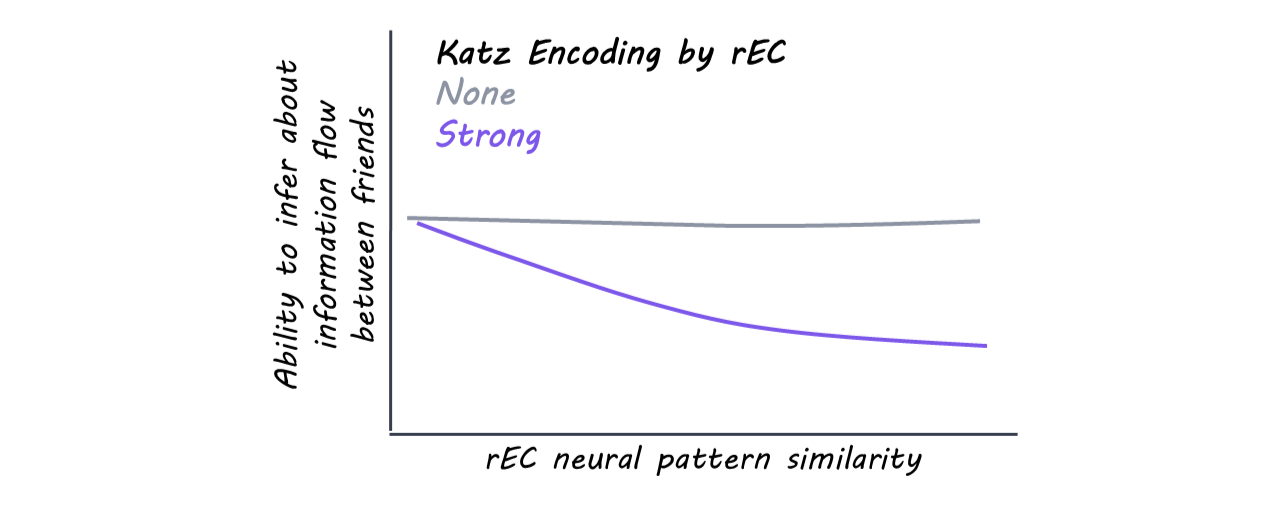
Finally, participants again relied on a Katz communicability-based strategy when reasoning about how news would spread through the network. Critically, when the right EC strongly encodes Katz communicability, the more strongly the right EC predicts task performance.
What's the impact?
This study is the first to demonstrate that the human brain encodes large-scale, real-world social networks as structured cognitive maps, within the MTL. These findings extend the MTL's known role in spatial navigation into the domain of complex social cognition. This work lays the foundation for investigating how social cognitive maps are updated over time, and how their disruption may contribute to deficits seen across neurological and psychiatric conditions.