Can Brain Stimulation Help Jump-Start Awareness?
Post by Annika Matthiesen
The takeaway
Stimulating specific brain regions may help “jump-start” brain activity in patients who have limited awareness and disrupted brain communication. While the effects were modest, the study suggests this noninvasive approach could help support recovery of awareness and responsiveness.
What’s the science?
People with severe disorders of consciousness have limited awareness and responsiveness because communication between different parts of the brain can be disrupted. Currently, there are very few effective treatments to improve wakefulness or recovery in these patients. This week in NeuroImage, Sangare and colleagues aim to improve brain activity and awareness in patients with limited wakefulness through brain stimulation.
How did they do it?
The authors studied 18 patients with disorders of consciousness. The authors used brain stimulation on the frontal cortex, an area at the front of the brain involved in higher-level thinking and control. Before and during treatment, the researchers measured how patients responded to sounds, sights, movement, and attempts to communicate. They also used EEG to record the brain’s electrical activity through sensors placed on the scalp, to track changes in brain function. Patients received stimulation or no stimulation (control), and the authors compared their responses before and after treatment. Six months later, the researchers followed up again to see whether any changes lasted over time.
What did they find?
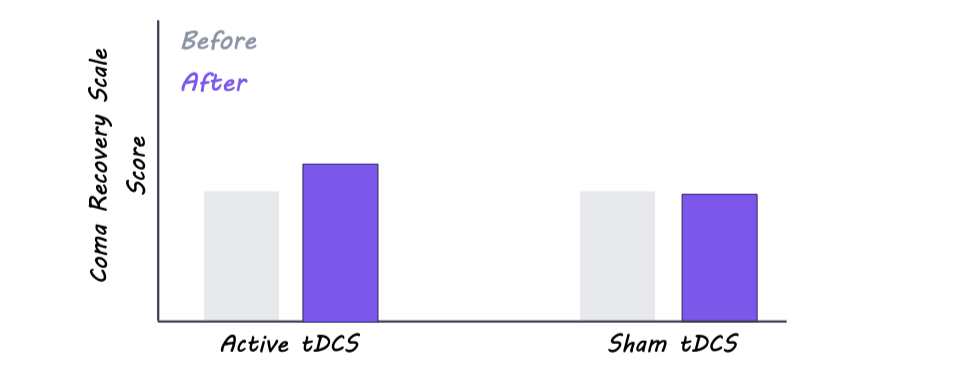
The researchers found some signs that the brain stimulation may have helped improve patients’ responsiveness and awareness, as the treatment group showed slightly greater behavioral improvement than the control group after stimulation. However, they did not observe changes in the overall level of consciousness. EEG results showed more active and complex brain activity in certain regions after treatment, which may suggest stronger communication between different parts of the brain. Overall, stimulation increased electrical activity across seven areas throughout the brain, although these changes did not consistently lead to major improvements in consciousness or behavior. Together, the findings suggest the stimulation may influence brain function, but larger studies are needed to determine whether it can produce meaningful recovery for patients with severe disorders of consciousness.
What’s the impact?
This study found that brain stimulation may help improve brain activity and responsiveness in patients with limited wakefulness by strengthening communication across different parts of the brain. If future studies confirm these findings, noninvasive brain stimulation could offer a safe way to improve awareness and brain communication in people with a wide array of serious neurological conditions.