How Bullying, Disordered Eating, and Depression Symptoms are Related
What's the science?
Adolescent bullying is a serious issue affecting approximately 30% of adolescents, and can have negative effects on mental health. In particular, bullying is associated with eating disorders and depression, but the longitudinal relationship between bullying, disordered eating and depression is not well understood. This week in JAMA Psychiatry, Lee and Vaillancourt tested the longitudinal relationship between these factors, in order to determine targets for intervention.
How did they do it?
The authors recruited adolescents aged 13-17, who were part of the McMaster Teen Study (a Canadian longitudinal study). 612 students completed questionnaires at several timepoints between ages 13 and 17 (a 5 year period). Students reported the frequency and type of bullying, to comprise a bullying severity score. The Short Screen for Eating Disorders questionnaire was used to measure the severity of disordered eating behaviour, and the Behaviour Assessment System for Children 2 was used to measure depression. The authors built several statistical models using these three variables plus control variables (sex, family income, race) to examine the relationships between these three factors, and assessed how well these models fit the data. In one model, ‘cross-lag’, meaning the effect of one factor at one timepoint on a different factor at a later timepoint, was modeled using a cascade modeling approach.
What did they find?
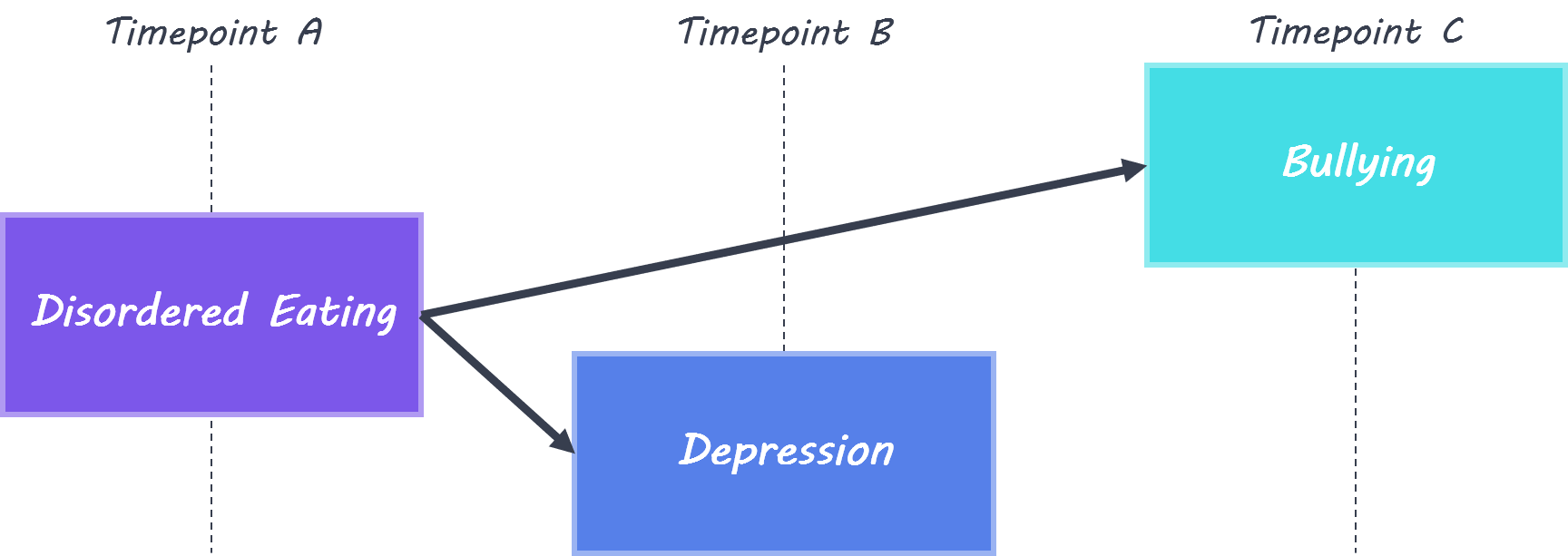
At each timepoint, there were significant positive relationships between the severity of bullying, disordered eating, and depressive symptoms. Girls reported more severe bullying, disordered eating, and depressive symptoms than boys. In the model which included ‘cross-lag’ effects, disordered eating was linked to depression one year later (at all timepoints) and to bullying two years later (at two timepoints). When sex differences were included in the model, it was discovered that there was a stronger relationship between disordered eating and depression in girls at most timepoints, while depression was more stable over time in boys.
What's the impact?
This is the first study to assess the relationship between bullying, disordered eating and depression in adolescents longitudinally. The results indicate that disordered eating at an earlier timepoint can predict both depression and bullying at later timepoints. This could be due to shared genetic factors underlying disordered eating, depression and bullying, or emotional dysregulation which is tied to disordered eating and could be a risk factor for bullying. This study suggests that targeting disordered eating and associated symptoms could reduce future depression and bullying in adolescents.

K.S. Lee and T. Vaillancourt, Longitudinal Associations Among Bullying by Peers, Disordered Eating Behavior, and Symptoms of Depression During Adolescence. JAMA Psychiatry (2018). Access the original scientific publication here.