
Understanding How Decisions Are Terminated
Post by Kulpreet Cheema
The takeaway
Little is known about how decisions are terminated and translated into actions or plans. This study provides evidence that the superior colliculus (SC), a midbrain structure involved in eye movements and orienting behaviors, plays a crucial role in terminating decisions.
What's the science?
Previous research has shed light on how the brain accumulates evidence before reaching a decision. This process can be modeled as a stochastic drift-diffusion process or bounded random walk. Neurons in the lateral intraparietal area (LIP) have been shown to accumulate noisy evidence during decisions. However, how this process is terminated and translated into a specific action is still unknown. The SC is known for its role in generating eye movements, and is directly coupled to the LIP; the LIP projects to the SC and the SC projects back to LIP via the thalamus. In a study published in Neuron, researchers investigated the role of SC in applying a decision threshold to the accumulation of evidence represented in the LIP.
How did they do it?
The researchers recorded simultaneous neural activity in the LIP and SC while monkeys performed a motion-discrimination task. In the motion discrimination task, the monkeys were trained to make eye movements based on the direction of a moving stimulus. Researchers used high-density multi-channel electrodes to capture the firing rates of functionally similar neurons in both areas during the decision process. To directly test the involvement of the SC in decision termination, researchers performed focal inactivation by temporarily inactivating the SC using small muscimol injections. They analyzed behavioral measures and neural recordings from the LIP during SC inactivation.
What did they find?
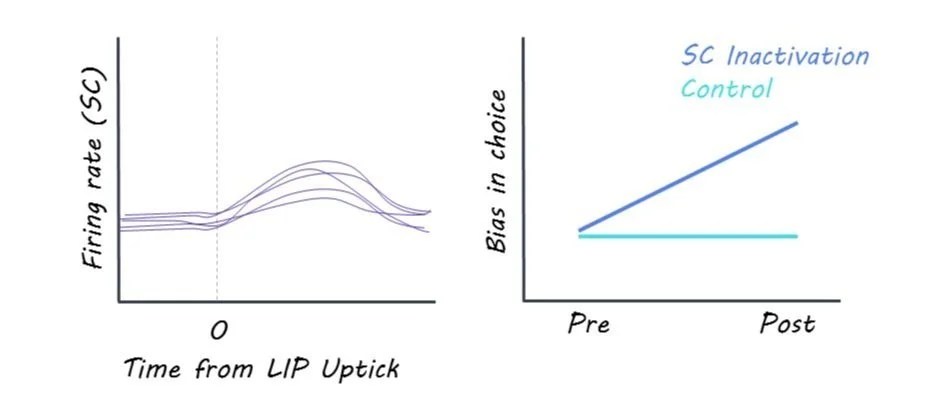
Researchers found evidence that the LIP and SC exhibited different dynamics during decision-making, and that the SC implements the decision threshold. The researchers observed that bursts of activity in the SC terminated the decision process, as opposed to the accumulation signals in LIP. The bursts in the SC were triggered by upticks in excitatory input and were associated with the termination of the decision. When the SC was inactivated, the termination mechanism was impaired, leading to slower, biased decisions and prolonged evidence accumulation in LIP. These findings suggest that, while evidence may accumulate in the LIP during decision-making, decision termination occurs in an area responsible for action selection; in this case, the SC, as the decision is about moving the eyes.
What's the impact?
This study sheds light on how the brain makes decisions and transforms evidence into actions. Understanding how decisions are terminated is crucial for comprehending the entire decision-making process. By identifying the role of the SC in decision termination, the study highlights the importance of a region known for action selection in decision termination. The findings have implications for understanding decision-making processes in humans and other primates.

